BIOCHEMISTRY TOPICS
Glycogen phosphorylase
Glycogen phosphorylase catalyzes glycogen degradation. Regulation of glycogen phosphorylation by phosphorylation and dephosphorylation, as well as allosteric effects. Reaction mechanism. Historical development of research on glycogen metabolism and its regulation.
Glycogen phosphorylase [EC 2.4.1.1],
often referred to simply as "phosphorylase" is the enzyme that catalyzes
the mobilization of glucose residues from storage in the polymeric form of glycogen.
The phosphorolysis of a terminal α-1,4 linkage (phosphate
is used to split the linkage) results in the production of glucose 1-phosphate
and a glycogen chain that is one glucose unit shorter.
Phosphorylase is tightly regulated in its activity by allosteric effectors and by phosphorylation.
It is also expressed as different tissue-specific isozymes.
Phosphorylase exists in three isoforms in mammals: liver, muscle, and neuronal.
The activity of muscle phosphorylase is allosterically regulated by ATP
(inhibited) and AMP (activated),
while the activity of liver phosphorylase is allosterically regulated (inhibited) by glucose.
Both isozymes are activated by phosphorylation
carried out by the kinase phosphorylase kinase
[EC 2.7.11.19].
The activity of phosphorylase kinase, is in turn regulated by cAMP-dependent protein kinase,
also called protein kinase A
(PKA) [EC 2.7.11.11],
a kinase that responds to the activation of a
hormone-responsive
signal transduction pathway involving heterotrimeric G-proteins and adenylate cyclase.
The hormones epinephrine and glucagon
turn on this pathway, raising levels of cAMP,
activating PKA, thus leading to the phosphorylation of phosphorylase kinase and phosphorylase.
The breakdown of glycogen is thereby accelerated.
Regulation of phosphorylase by reversible phosphorylation is diagrammed below at left.
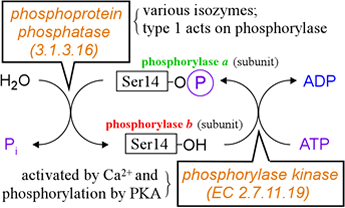
A serine residue near the N-terminus of the enzyme (Ser14) is targeted by phosphorylase kinase. Attachment of a phosphoryl group to Ser14 converts the intrinsically less active form, called phosphorylase b, to the intrinsically more active form, phosphorylase a. The "a" in this context can be interpreted as meaning "active".) The reverse process, removal of the phosphoryl group from Ser14, is catalyzed by a phosphatase, phosphoprotein phosphatase I, which is one of a number of protein phosphatase [EC 3.1.3.16] isozymes.
The phosphatase and kinase that catalyze the interconversion of phosphorylase a and b are themselves regulated, and this is also shown in the figure. The kinase is activated by calcium ion (Ca2+) and phosphorylation by protein kinase A (PKA).
Mechanism of glycogen phosphorylase
The mechanism of phosphorylase prominently features general acid - base catalysis. The active site contains a pyridoxal phosphate (PLP) cofactor, which acts in the mechanism as a general acid and a general base. This is an atypical role for PLP, which - derived from vitamin B6 - is most notable for its catalysis in transformations of amines and amino acids. Here, PLP donates a proton to substrate Pi as the latter in turn donates a proton to the leaving 4′ oxygen forming the new nonreducing end of the glycogen chain, as an oxonium intermediate of the cleaved glucose unit is generated. Pi then adds to the re face of the planar oxonium to form glucose 1-phosphate with retention of the α configuration.
Historical development of research on glycogen metabolism
In the 1940s, Carl and Gerty Cori discovered phosphorylase, showing it to be the enzyme that degrades glycogen to glucose 1-phosphate (the latter, which they had isolated from minced frog muscle, was historically called the "Cori ester"). Furthermore, the Coris found that phosphorylase activity could be affected by small molecules (AMP, ATP, glucose) and that phosphorylase could be purified in two different stable forms. One - called phosphorylase a - was active without addition of AMP. The other, termed phosphorylase b, was inactive unless AMP was present. This was the first description of what we now recognize as allosteric regulation of an enzyme's activity. The Coris had even identified an enzyme they dubbed "PR" for "prosthetic group-removing" enzyme, which converted phosphorylase a to phosphorylase b.
It was not until the 1950s, when research - principally by Edwin Krebs (who had trained with the Coris) and Edmond Fischer - elucidated the novel mechanism of regulation of glycogen phosphorylase by reversible phosphorylation. They found that a kinase, phosphorylase kinase [EC 2.7.11.19], phosphorylates the enzyme, converting it to the a form, and showed that the Cori's PR was a protein phosphatase [EC 3.1.3.16]. Fischer and Krebs won the 1992 Nobel Prize for Physiology or Medicine for their discoveries of "reversible protein phosphorylation as a biological regulatory mechanism".
Glycogen phosphorylase structure, function and regulation
Regulation of phosphorylase activity occurs by phophorylation and dephosphorylation, as well as by allosteric effects. Furthermore, isozymes of phosphorylase that are characteristic of different organs in mammals differ in their predominant allosteric regulatory features. Comparing the liver and skeletal muscle isozymes illustrates how regulation can be adapted to tissue-specific roles.