BIOCHEMISTRY TOPICS
ATCase: "the hemoglobin of enzymology"
Aspartate transcarbamoylase (ATCase): Structure, role as an enzyme in a biosynthetic pathway and in regulation by feedback inhibition. Allosteric properties of ATCase.
Aspartate transcarbamoylase [EC 2.1.3.2], which catalyzes the committed step in the pathway for pyrimidine nucleotide synthesis in bacteria, is dubbed "the hemoglobin of enzymology" since it is the most well-studied allosteric enzyme. The substrates of the ATCase reaction are carbamoyl phosphate (CbmP) and aspartate; the products are N-carbamoylaspartate plus orthophosphate.
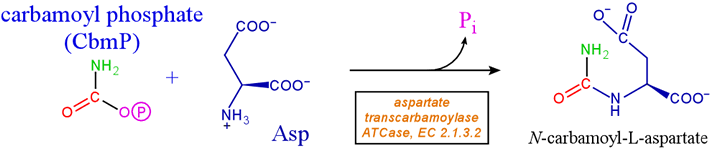
The following step in the pathway is cyclization of N-carbamoylaspartate to dihydroorotate. Gerhart and Pardee found in 1962 that the enzyme displays sigmoidal kinetics, meaning the reaction velocity vs. substrate concentration curve has a sigmoidal form. The enzyme shows a strong preference for binding of CbmP first. Once CbmP is bound, the affinity of the enzyme for aspartate is increased markedly, and aspartate shows a positive cooperative effect on catalysis.
The reaction likely proceeds through a tetrahedral transition state after the α-amino group of aspartate attacks the carbonyl carbon of carbamoyl phosphate, and prior to the expulsion of the orthophosphate leaving group. Note that CbmP is a fairly high-energy phosphoric anhydride, making this reaction energetically favorable (energy is required to synthesize CbmP).
The first structure of ATCase was determined by X-ray crystallography in 1982 by William Lipscomb and associates (2.6 Å resolution). It showed a dodecameric (12 subunit) quaternary structure constructed of two catalytic trimers separated along a vertical axis corresponding to a three-fold symmetry axis, with the catalytic trimers bridged by three regulatory dimers.
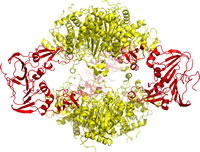
This dodecameric structure [see 6at1.pdb], referred to as the holoenzyme (to distinguish it from catalytic trimers and regulatory dimers), was fully consistent with biochemical studies of ATCase. Catalytic trimers (in yellow in figure at right) and regulatory dimers (red) can be isolated from intact holoenzyme by treatment with mercurial reagents followed by separatory procedures. The catalytic trimers have full catalytic activity and show no cooperative behavior.
The role of the regulatory subunits of ATCase is to mediate heterotropic effects, analogous to the Bohr effect or that of 2,3-BPG in hemoglobin. Cytidine triphosphate (CTP), which is an end product of the pyrimidine biosynthetic pathway, has a negative allosteric effect on ATCase activity, while adenosine triphosphate, ATP, has a positive allosteric effect. CTP and ATP compete for the same binding sites within each regulatory subunit. The negative allosteric effect of CTP is an example of feedback inhibition, a typical mechanism by which biosynthetic pathways are regulated.
An extremely useful inhibitor of ATCase known as PALA is a bisubstrate analog. PALA is a non-hydrolyzable phosphonate compound which is similar to CbmP + aspartate, except for a missing amino group.
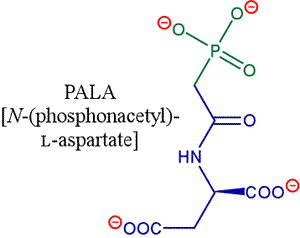
PALA has a Kd of about 10 nM for ATCase, meaning it binds the enzyme quite strongly. The crystal structure of ATCase in complex with PALA [1d09.pdb] was used not only to infer the nature of enzyme-substrate contacts, but also (by comparison to the structure of the free enzyme) in the effort to define the nature of the structural changes accompanying allosteric effects.
The location of PALA in the structure of ATCase with bound PALA shows that the active site is formed at the interface between subunits within a catalytic trimer. Comparison of the structures of unliganded ATCase holoenzyme and PALA-bound ATCase holoenzyme reveals striking differences. The latter shows an increase in separation of the catalytic trimers by 12 Å, as well as a relative rotation of 10º. This dramatically reduces the interactions between c chains in different opposing trimer. The regulatory chains also rotate, accommodating the larger distance between the catalytic trimers. The individual c chains within the catalytic trimer also undergo a structural transition. The two domains within the c subunit approach one another more closely by a hinge motion of about 8º, resulting in a closure at the active site of about 2 Å. The closure of the active site is stabilized by interdomain bridging interactions.
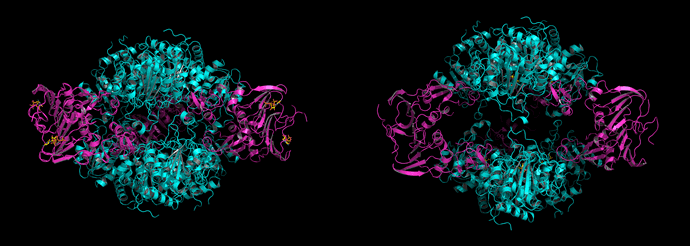
Above: The structures, represented in ribbon form, of ATCase holoenzyme with CTP bound (left), and with PALA bound (right). In both cases, the catalytic trimers are shown in cyan, and the regulatory dimers are colored magenta. Note that the expansion of the PALA-bound structure results in the nearly complete separation of catalytic trimers, as opposed to the structure on the left in which there are extensive contacts between the trimers. The ligands (CTP and PALA, respectively) are shown as sticks (carbon atoms are colored yellow). CTP binds stoichiometrically to the regulatory subunits, and is readily visible in the structure at left. PALA binds to each of the three active sites formed by each catalytic trimer. The figure on the left is based on 1rab.pdb - and the figure on the right is based on 1d09.pdb. Both the images were generated using PyMOL (Ref.8).
The structural information and the kinetics studies of ATCase suggest that its behavior may be well modeled by the MWC (Monod-Wyman-Changeux) or symmetry model. If the structure of unliganded ATCase holoenzyme or the CTP-bound form represent the T state, and the PALA-bound ATCase holoenzyme represents an R state, then the structural changes described above correspond to a conformational equilibrium between T and R states of the holoenzyme. The increased separation of catalytic trimers results in a deconstraint of the c subunits and allows the hinge movement between domains by which the active site attains its optimal structure and activity. The tertiary and quaternary changes in ATCase are tightly coupled, and in many respects the two-state model works especially well in this case.
There is a twofold biological significance of the regulatory features of ATCase. Feedback inhibition by CTP and activation by ATP tends to coordinate the rates of synthesis of purine and pyrimidine nucleotides. Such synthesis should occur in the cell only when energy (in the form of ATP) is available.